Rev Bras Fisiol Exerc 2020;19(2);154-71
REVIEW
Metabolism
of fatty acids, secondary complications and effects of physical exercise:
integrative review
Metabolismo dos ácidos
graxos, complicações secundárias e efeitos do exercício físico: revisão
integrativa
Djeyne Silveira Wagmacker1,2,
Alice Miranda de Oliveira3, Edna Conceição de Oliveira2,
Alan Carlos Nery dos Santos1,4, Luiz Erlon
Araújo Rodrigues1, Ana Marice Teixeira
Ladeia1
1Escola Bahiana de Medicina e Saúde Pública, Salvador, BA, Brasil
2Faculdade Adventista da
Bahia, Cachoeira, BA, Brasil
3Centro Universitário
Social da Bahia, Salvador, BA, Brasil
4Grupo de Pesquisa
Ciências da Saúde em Fisioterapia, Universidade Salvador - UNIFACS, Feira de
Santana, BA, Brasil
Received on
March 15, 2020; accepted, April 5, 2020.
Corresponding author: Alice Miranda de
Oliveira, Centro Universitário Social da Bahia, Av. Oceânica, 2717, Ondina,
40170-010, Salvador, BA
Djeyne Silveira Wagmacker: djeyne.ferreira@adventista.edu.br
Alice Miranda de
Oliveira: alicemofisio@gmail.com
Edna Conceição de
Oliveira: ednamoreira1868@gmail.com
Alan Carlos Nery dos
Santos: alancarlos.nery@unifacs.br
Luiz Erlon
Araújo Rodrigues: erlon@ufba.br
Ana Marice
Teixeira Ladeia: analadeia@uol.com.br
Abstract
Introduction: Diet is a complex set of exposures that frequently interact, and whose
cumulative effects influence the results of health. This includes effects on
systemic inflammation markers in metabolic disturbances and cardiovascular
diseases. Various studies have been presented relating the effect of physical
exercise on lipids, however, the results are still controversial. Objective:
To describe fatty acid metabolism and the effect of physical exercise on
secondary complications. Methods: An integrative review was conducted on
topics in the Medline, Pubmed, Web of Science and
Scopus databases, published up to the year 2017. Results: Fatty acids,
depending on their biochemical characteristics and spatial configuration, have
differentiated effect on cardiovascular health, however, studies still present
contradictory results about the therapeutic use of certain fatty acids.
Physical exercise appears to benefit fatty acid metabolism and attenuate the
complications secondary to the intake of certain fatty acids, and potentializes
the positive effects of distinct fatty acids. Conclusion: However,
variants of physical exercise, such as intensity, duration, time of observation
of effects of the results, limit the authors to concluding, with a certain
degree of certainty, about the effect of physical exercise on fatty acids and
secondary complications, since the studies in the literature continue to be
contradictory.
Keywords: fatty acids; exercise; inflammation; oxidative stress.
Resumo
Introdução: A dieta é um conjunto
complexo de exposições que interagem frequentemente e cujos efeitos cumulativos
influenciam os resultados da saúde. Isso inclui efeitos nos marcadores de
inflamação sistêmica em distúrbios metabólicos e doenças cardiovasculares.
Vários estudos foram apresentados relacionando o efeito do exercício físico
sobre lipídios, no entanto, os resultados ainda são controversos. Objetivo:
Descrever o metabolismo dos ácidos graxos e o efeito do exercício físico nas
complicações secundárias. Métodos: Foi realizada uma revisão integrativa
dos assuntos nas bases de dados Medline, Pubmed, Web of Science e Scopus, publicadas até o ano de 2017. Resultados:
Os ácidos graxos, dependendo de suas características bioquímicas e configuração
espacial, têm efeito diferenciado na saúde cardiovascular, no entanto, estudos
ainda apresentam resultados contraditórios sobre o uso terapêutico de certos
ácidos graxos. O exercício físico parece beneficiar o metabolismo dos ácidos
graxos e atenuar as complicações secundárias ao consumo de certos ácidos
graxos, além de potencializar os efeitos positivos de diferentes ácidos graxos.
Conclusão: No entanto, variantes do exercício físico, como intensidade,
duração, tempo de observação dos efeitos dos resultados, limitam os autores a
concluir, com certo grau de certeza, sobre o efeito do exercício físico sobre
ácidos graxos e complicações secundárias, uma vez que estudos na literatura
continuam sendo contraditórios.
Palavras-chave: ácidos graxos;
exercício; inflamação; estresse oxidativo.
Introduction
Cardiovascular diseases continue to be the main cause of morbidity and
mortality in the world, despite improvements in the results [1,2]. However,
risk factors such as obesity and diabetes mellitus (DM) have increased
substantially and increased the inequalities among countries. Not only the
prevalent risk factor cause concern about these diseases, but also the low
level of implementation of preventive measures, such as low-quality diet and
physical inactivity [3].
A large portion of the cardiovascular disturbances have their origin in
atherosclerosis, characterized by changes in the intima, represented by
accumulation of lipids, components of the blood, cells, intercellular matter
and carbohydrates [4].
Lipids have always been present in diets. Diet is a complex set of
exposures that frequently interact, and whose cumulative effects influence the
results of health. This includes effects on systemic inflammation markers in
metabolic disturbances and cardiovascular diseases [5].
The fat consumed is composed of fatty acids (FA) and glycerol. The
larger part of FAs in humans are of the long chain type, divided into saturated
and unsaturated types that may present a cis or trans configuration [6]. The
composition of FAs coming from the diet is an important factor, because they
cause different metabolic changes [7].
At present, an increase in trans-fat consumption by individuals has been
observed. This has aroused the interest of the scientific community, because
the consumption of trans fatty acids has been related to increased risk of
coronary diseases [8], changes in plasma lipoproteins and triglycerides,
increased risk for Diabetes Mellitus [9], elevation of serum inflammatory
markers [5], oxidative stress and endothelial dysfunction markers, as well as
worsened nitric oxide-mediated vasodilator response [10]. The physical-chemical
characteristics of fatty acids, such a melting point, carbon chain size,
presence of double bonds and geometric configurations are important aspects
that may interfere in the absorption of fatty acids by tissues, especially the
adipose and vascular types, and the development of health problems.
Various studies have been presented relating the effect of physical
exercise on lipids, however, the results are still controversial [11-14]. Over
the last few decades it has been possible to observe growing evidence that
acute physical exercise could have an acute beneficial influence on the lipid
profile [15,16]. The difficulty with analyses and interpretation of these
studies lies in the use of different physical activity protocols established.
Fatty
acids: definition and classification
Lipids are distinct elements among them, presenting different chemical
and functional characteristics. Fatty acids, the constituent elements of
lipids, are present in any lipid structure.
Fatty acids are organic components that contain carbon and hydrogen in
their molecules. Depending on the type of combination among fatty acids and
their constituents, they will form different types of lipids. Based on these
combinations, they may be classified as simple or complex types. Simple lipids
are those in which the fatty acid combines with only one other element (e.g.:
triglycerides), whereas, complex lipids are those in which the fatty acid
combines with more than one element (e.g.: lipoproteins) [17].
Among the characteristics that distinguish the fatty acids is the size
of the carbonated chain. Fatty acids may also be classified as short and long
chain types. The short chain type has between 4 and 16 carbon molecules, and
when they are not supplied by the diet, they are synthesized, mainly in the
cytoplasm of hepatic and adipose tissue cells. The long chain fatty acids have
16 or more carbon molecules, and when they are not supplied by the diet, they
are formed by elongation of pre-existent fatty acids [18]. In addition, fatty
acids may be classified based on the type of bond among their molecules,
differing between saturated or unsaturated types. Unsaturated fatty acids have
double bonds between their carbon molecules, while the unsaturated type does
not have these bonds [19]. Unsaturated fatty acids are chemically more unstable
and may be of the monounsaturated type, when they have only one double bond, or
polyunsaturated when they have two or more double bonds [20].
Unsaturated fatty acids may present a cis or trans configuration. They
are characterized as a cis fatty acid when the hydrogen molecules in their
geometric configuration are presented in the same plane as that of the double
carbon bond. They are characterized as an unsaturated fatty acid of the trans
type when their hydrogen molecules are on the opposite side of the double
carbon bond. The trans fatty acid is a geometric isomer of the original cis
fatty acid; that is, it presents the same quantity of carbon, oxygen and
hydrogen molecules but with a different spatial configuration [19].
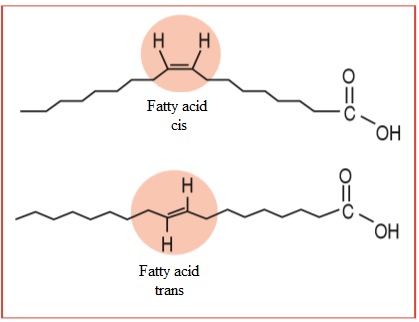
Figure
1 – Illustration of o cis-unsaturatel
fatty acid and a trans-unsaturated fatty acid. Source: Lima [21]
The two types of fatty acids may be found in nature, however, the cis
configuration is more common, because the enzymes that synthesize fatty acids
prefer this configuration. The trans fatty acids found in nature are present,
in a reduced manner, in meats and milk. More expressive incorporation of trans
acids into the human diet occurred with the process of hydrogenation of
vegetable oils, especially by the food industry. Heating vegetable oil also
induces the formation of geometric isomers of polyunsaturated fatty acids, and
so does the irradiation of foods [22].
Trans fatty acids are more stable than the cis fatty acids, and
therefore less energetic. Because the cis isomers are more energetic, they
would be involved in the synthesis of different cellular lipids. The change in
the structure of fatty acids to the trans form
modifies the melting point, increases the plasticity and oxidative stability of
these fats. Elaidic acid, for example, (9trans-18:1)
presents a melting point of 440C whereas, oleic acid (9cis-18:1) has a melting
point of 130C [23].
Melting point is an important characteristic of fatty acids. The higher
the melting point, the greater the quantity of thermal energy necessary to
break down their molecular arrangements. This allows greater impregnation into
the tissues such as the vascular endothelium, and particularly in the adipose
tissue. As the carbonated chain length increases, the melting point also
increases. However, the presence of double bonds makes the melting point fall. Due
to the geometric configuration, the melting point of trans fatty acids is also
higher [24].
When trans fatty acids are ingested and absorbed, they may change the
composition and biochemical activity of the cell membranes, thus changing the
physical properties of the membrane, and finally change its functions [20,24].
The composition of phospholipids in the plasma membrane has a crucial influence
on cellular growth and metabolic activity. In the last two decades, studies
have suggested that the lipid composition in the diet influences the fatty acid
profile of serum and the lipid content of the plasma membrane. In fact, the
length of the fatty acid chains and the degree of saturation or unsaturation
have been shown to change the fluidity and activity of various proteins
associated with the membrane [25].
With the change in the physiological processes because of the
incorporation of these fatty acids into the different tissues, evidence of
different adverse clinical situations has been shown in the literature. Among
these, for example, increase in triglycerides, change in lipoproteins such as
increase in LDL, VLDL, reduction in HDL concentration [26], increase in insulin
resistance, increasing the risk of diabetes Mellitus [27], increase in the production
of pro-thrombotic factors, increase in reactive oxygen species [28], increased
risk for CAD, especially Acute Myocardial Infarction [29], among other
pathologies [23].
Studies such as that of Chajés et al.
[30], have suggested that high ingestion of industrial trans fatty acids could
cause an increase in body weight, especially in women. Furthermore, as
mechanism for the prevention of obesity, they have suggested limitation of the
consumption of highly processed foods must be considered, as they are the main
source of industrially produced trans fatty acids [31].
On the other hand, unsaturated fatty acids with cis configuration have
been related to favorable effects on the metabolism. Evidence has been shown of
association between the consumption of unsaturated fat and aspects such as
reduction in: blood viscosity and plasma
triglycerides; higher level of endothelium relaxation [32]; improvement in
insulin sensitivity [33], among others.
Evidence has been shown that the intermediate products of fatty acid
metabolism are important for myoblast survival, proliferation, differentiation
and fusion. Studies have suggested that the lipid metabolites derived from
polyunsaturated fatty acids accelerated protein synthesis, and the fusion and
growth of muscle cells in different animal models [34].
Fostok et al.
[35], have demonstrated that oleic acid (cis, unsaturated fatty acid)
supplementation attenuated incomplete repair actions, optimizing the
regenerative capacity and contractile function of the injured muscle.
Despite the body of evidence presented up to now, substitution of
saturated fats with polyunsaturated fats in diets as a way of preventing
cardiovascular diseases continues to be questionable. According to Hamley [36], in a meta-analysis published in 2017, the
evidences available in the randomized clinical trials up to now suggested that
the substitution of saturated fatty acids with polyunsaturated fatty acids
(n-6) in diets was not sufficient to reduce the events of cardiovascular diseases,
mortality due to coronary disease or total mortality. Furthermore, he suggested
that his results have implications for present dietary counsel, in which the
recommendations to reduce saturated fat and/or substitute saturated fatty acids
with polyunsaturated fatty acids must not be emphasized, because the
maintenance of these recommendations would probably not have the intended
effect and could reduce the efforts towards (encouraging) persons to adopt
other lifestyle changes that would most probably be more beneficial.
The chart below presents the summary of the studies cited above, on the
effect of fatty acids on the cardiovascular and metabolic systems.
Frame
1 - Summary of the studies about fatty acids and
effect on the cardiovascular and metabolic systems
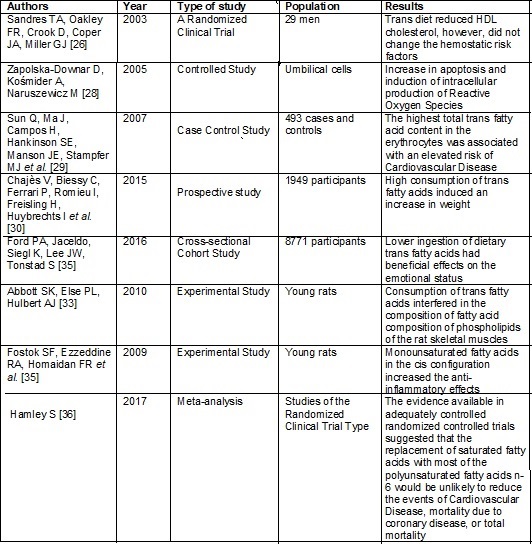
Fatty
acid absorption and metabolism
Approximately 90% of the lipids consumed are in the form of
triglycerides (TG). The remaining 10% are in the form of cholesterol,
cholesterol esters, phospholipids and free fatty acids. Lipid digestion begins
in the mouth and stomach with the action of the lingual and gastric lipases.
They degrade the TGs into medium and short chain fatty acids. After the action
of the lipases, they undergo the action of the biliary salts and substances
produced by the pancreas such as pancreatic lipase that will degrade the TGs
formed by long chain fatty acids [37].
In sequence, the short and medium chain free fatty acids are directly
absorbed by the enterocytes of the intestinal mucosa and are released into the
venous blood stream. They are then transported to the liver by albumin, by the
hepatic port circulation, or to the peripheral tissues in which they are
directly absorbed and used as energy substrate. Whereas, the long chain free
fatty acids, non-esterified cholesterol, phospholipids together with their
biliary salts and the liposoluble vitamins (A, D E and K), form the mycelia,
which are hydrophilic particles that facilitate lipid transport, and these
liposoluble vitamins, through the membrane of the enterocytes [38].
After crossing the intestinal mucosa, the compounds of the mycelia,
together with Apolipoprotein B-48, will form the primogenitor lipoprotein – Chylomicra or Chylomicrons. At this point, the fatty acids
are again resynthesized into triglycerides and the free cholesterol is
esterified. Therefore, the chylomicras mainly
transport triglycerides and esterified cholesterol [39].
The chylomicras are then transported to the
peripheral tissues, to which mainly fatty acids are released for energy production.
To enable them to be released, these fatty acids must be cleaved from the
glycol. Lipoprotein lipase is the enzyme that cleaves the triglycerides coming
from the chylomicras and later from the VLDL. The
free fatty acids are then absorbed by the adipose or muscle tissue, or then
transported to other tissues by albumin [37].
Lypolysis
comprises four stages: cleavage of the triglycerides in the blood;
beta-oxidation of the fatty acids; the citric acid cycle, and the electron
transport chain. The first stage, as previously described, occurs in the blood,
where the triglycerides present in the low-density lipoproteins are cleaved by
lipoprotein lipase. In this process, the fatty acids are released from the
glycerol and transported up to the sarcolema (plasma
membrane of the muscle cell) by albumin. On entering the cell, the second stage
- beta-oxidation - occurs. In this stage, the fatty acids undergo the action of
enzymes, such as thiokinase, present in the external
membrane of the mitochondria. The mitochondria have two membranes - internal
and external - and a space between these membranes. Beta-oxidation is finalized
in the external membrane of the mitochondria, the site where thiokinase is found, which will finalize this process.
However, the long chain fatty acids, differently from the medium and short
chain types, are not permeable to the internal membrane of the mitochondria,
and requires the action of carnitine, nutrient transporter of AcetylCoA, resulting from the catabolism of the long chain
fatty acids through the internal membrane of the mitochondria [40].
Oxidative metabolism allows energy to be obtained from fatty acids in an
intramitochondrial localization. Thus, to enable acyl-CoA to be used by it
(oxidative metabolism), it is necessary to overcome the impermeability of the
external and cytoplasmic membrane of the mitochondria to attain the acyl-CoA.
The enzyme responsible for this transport is Carnitine-CoA acyltransferase
(Carnitine O-Palmitoyltransferase). This enzyme
presents greater specificity for Palmitoyl-CoA,
however, it catalyzes transport of fatty acids with a carbonated chain length
between C4 and C18. Fatty acid chains longer than these are more difficult to
be transported. Once inside the mitochondria, acyl-CoA may be used in the
lipolytic metabolism of LYNEN [41].
Carnitine palmityl transferase was historically seen as the only
regulator of fatty acid oxidation. However, other FA translocators, such as
FAT/CD36 have been identified. Specifically, FAT/CD36 appears to have a
differentiated mechanism of action with respect to fatty acid oxidation during
exercise, influencing lipid transport through the sarcolemal
membrane and to the mitochondria [42].
Differently from the striated muscles, adipose tissue allows the entry and
exit pathway of fatty acids. While the fatty acids only enter the muscles with
the purpose of producing energy, whereas, on entering the adipose tissue, the
fatty acids may produce energy and may also be accumulated. When necessary, the
fatty acids accumulated in the form of triglycerides may be hydrolyzed and thus
release fatty acids into the blood stream, and these are transported by albumin
to be used in other tissues to produce energy. The main consumer of these fatty
acids released by the adipose tissue are the striated cardiac and skeletal
muscles [40].
Fatty
acid metabolism during exercise
The process of fatty acid uptake and oxidation is important for ATP resynthesis [43]. Once the fatty acids enter the skeletal
fiber, they have different destinations, depending on the metabolic state of
the cells. In conditions of rest, the plasma fatty acids are conducted to
triglyceride synthesis as the first destination, instead of being moved to the
mitochondria for oxidation [43].
As the exercise progresses, long chain fatty acids, provided by the
blood or from hydrolysis of intramuscular triacylglycerols, are metabolized to
generate energy. The supply of fatty acids from the hydrolysis of intramuscular
triacylglycerols is limited and during exercise; the myocytes consume
approximately 90% of the free fatty acids derived from blood plasma [41].
Small quantities of the triglycerides are stored within the lipid
droplets in the skeletal muscle and may be hydrolyzed to produce fatty acids
for energy production by means of b oxidation and oxidative phosphorylation.
Although there has been some controversy about the quantitative importance of
intramyocellular (IMTG) as metabolic substrate, recent studies have
demonstrated a substantial contribution by IMTG to energy production [45].
There are three lipases expressed in the skeletal muscle, which are
responsible for the degradation of TG: monoacylglycerol lipase; Adipose
triglyceride lipase (ATGL) and hormone-sensitive lipase (HSL).
ATGL is the first step of TG lipolysis in the skeletal muscle of humans
and mice, resulting in the release of one fatty acid molecule. Monoacylglycerol
lipase is responsible for the hydrolysis of monoacylglycerol, releasing
glycerol and fatty acids. The HSL catalyzes the hydrolysis of TG to release FA
in the cytoplasm [41]. The HSL is highly present in the type I oxidative fibers
of skeletal muscle and is activated by adrenergic stimulation and contraction.
ATGL is activated by the comparative identification of genes-58 (CGI-58), These
proteins are localized on the surface of the mitochondria, and are preferentially
expressed in oxidative muscle, such as the cardiac and soleus muscles [45].
Resistance training leads to increased levels of ATGL, increasing intramuscular
lypolysis, particularly in Type I oxidative fibers
[46]. Studies such as that of Roepstorff et al. [47] have demonstrated
that exercise triggered the rapid activation of HSL dependent on protein
kinases in human beings, promoting the release of FA.
The mechanisms that regulate exercise-induced lipolysis in the skeletal
muscle have not yet been completely elucidated and may be more complex than
lipolysis in adipose tissue [48].
Studies have also suggested that physical exercise performed in an acute
manner promotes changes in the fatty acid transport genes, preceding increases
in RNAm expression [49]. This will allow greater
metabolization of fatty acids, although the latter possibility has not yet been
tested [50].
According to the research of Kim et al. [51], physical exercise is also
capable of significantly increasing the expression of components of the
metabolic pathway and components related to the redox signal induced a similar
increase in the FAT/CD36 content of the cell membrane of skeletal muscle in
rats.
In a randomized clinical trial published in 2017, the response of
post-prandial triglycerides was observed to be attenuated by low to moderate
intensity periodized exercises, when measured after 24 hours [52].
The use of lipids is modulated by the availability of fatty acids in the
plasma. In obese subjects, the plasma AG levels are more elevated. Glycolytic
activity is altered in this population, and lipid metabolism may be a
preferential route. In obese subjects, the metabolic responses of fatty acid
mobilization appear to be less favored by aerobic activity, however, the responses
are not yet conclusive [11].
Recent studies have reinforced the differences in the patterns of
lipolysis stimulation between thin and obese subjects during physical exercise.
The difference in the lipolytic rate appear to be due to differences in the
quantity or activity of the lipases present in the skeletal muscle, especially
ATGL, and not the mRNA levels [53].
Frame
2 - Effect of exercise on fatty acid metabolism
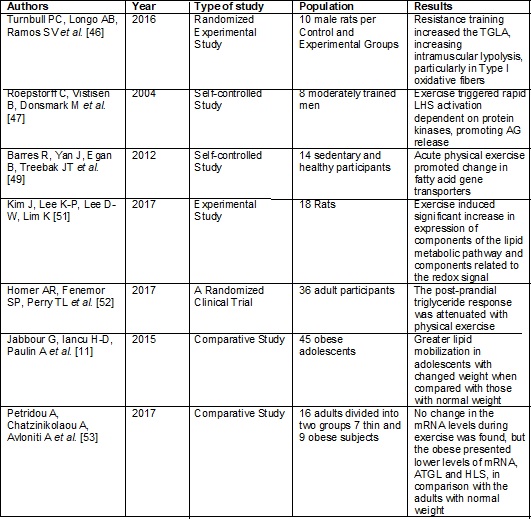
Fatty
acid consumption, inflammatory and endothelial dysfunction status
There are various mechanisms by means of which diet increases or
diminishes the risk for cardiovascular diseases. Investigation of the
mechanisms that determine atherosclerosis have suggested that an inflammatory
process plays a central role in its development, progression and outcomes. This
inflammatory process causes structural and functional changes in the blood
vessel walls, which leads to endothelial dysfunction and the development of
atherosclerotic lesions [54]. Calorie-restriction diets are known to reduce the
circulating levels of C-reactive protein, which is a systemic inflammation
marker that may also play its own role in the inflammatory process, and many
studies have shown that it prevented cardiovascular events [55]. Diets rich in
omega-3 fatty acids appear to reduce atherosclerosis by means of the process of
down-regulation of intracellular mechanisms that lead to the expression of
pro-atherogenic genes [5].
The vascular endothelium is considered a dynamic tissue, an “organ”
controlling important functions, such as coagulation, maintenance of blood
circulation, vagal tonus, fluidification and inflammatory responses. Among
these various functions, the endothelium is also responsible to produce
vasodilator and vasoconstrictive substances. Nitric oxide is the main factor in
dilator responses and is directly involved in endothelial dysfunction [56].
The term “endothelial dysfunction” refers to an imbalance in endothelial
production of mediators that regulate vascular tonus, platelet aggregation, coagulation
and fibrinolysis. Endothelial dysfunction is also frequently reported as
worsening in endothelium-dependent relaxation, caused by loss of nitric acid
(NO) bioavailability, although changes in other vasoactive substances have also
been found [57].
Nitric oxide has diverse antiatherogenic functions, among them,
inhibition of smooth muscle cell production, inhibition of platelet aggregation
and antioxidant properties. Its release is stimulated by the shear force exerted on the endothelium by blood; this fact
is shown by the higher level of NO released from the arteries, in comparison
with veins [56].
In the post-prandial state, a longer period during which
triglyceride-rich (TG) lipoprotein levels remain elevated, may lead to
endothelial dysfunction. This results in increase in the inflammatory response,
lower level of nitric acid availability, and increase in oxidative stress -
changes involved in the genesis of atherosclerosis [58].
After a meal with a high fat content, healthy individuals present a
significant increase in the concentrations of proinflammatory cytokines, TNF-α,
IL-6 and adhesion molecules (intercellular adhesion molecule-1 - ICAM-1 and
Vascular cellular adhesion molecule-1 - VCAM-1), when compared with a meal with
a high carbohydrate content. These changes may also be prevented with the use
of Vitamin E, suggesting that oxidative stress regulates the increase in
cytokines and adhesion molecules. Studies have demonstrated that the
post-prandial triglyceride levels, and not those coming from adipocytes in
fasting are more sensitive markers of atherosclerosis [59,60].
Elevation of C Reactive Protein (CRP) begins around 6h after
inflammatory stimulation; it has a half-life of approximately 19h, and its
maximum value is attained in 24-72h. Its plasma concentration is constantly
low, and does not present circadian variations, in contrast with some
coagulation proteins and others of the acute inflammatory stage. Once
stimulation has been concluded, the values return to normal after 7 days.
Exercise,
inflammatory response and endothelial dysfunction
Although studies have shown evidence that the practice of physical
activity prevents the genesis and progression of atherosclerotic disease, the
mechanisms to explain this effect have not yet been completely elucidated. Ghisi et al. [56] suggested that the factor
responsible for this effect is related to the change in vascular tonus and
endothelial function.
Atherosclerosis development and progression partly depend on the
migration of monocytes to the blood vessels, to become active and begin the
release of cytokines. According to Vuorimaa et al.
[61] the first cytokines in the cascade are the tumor necrosis factor (TNF-α)
and interleukin1 (IL1) considered proinflammatory cytokines. After an acute
exercise session, there is no increase in the proinflammatory cytokines,
suggesting that physical activity suppresses the entry of these cytokines into
the plasma.
Paton et al. [62], after a study with healthy and sedentary
subjects, concluded that exercise performed at 50% to 70% intensity
continuously for 6 months could improve the inflammatory response, coagulation
and fibrinolytic potentials, reducing the risk for cardiovascular disease.
The effects of physical exercise on endothelial function, has been
demonstrated in animal and human experiments, however, the literature is still
controversial relative to the intensity of effort necessary to cause protective
effects. The intensity most tested in both humans and animals is the moderate
level, however, there are some evidences that high intensity, acute aerobic
exercise increased the chance of cardiovascular events, but when performed
chronically, it was associated with decreased occurrence of these events and
mortality [58].
MacEneaney
et al. [63] in a study about the effect of post-prandial lipemia on the
inflammatory markers and endothelial activation in adolescents, found that
physical exercise did not change the values of C-Reactive protein, TNF-α,
IL-6 and adherence molecules in circulation, showing that in spite of
significant reductions in hyperlipemia, exercise did not change the
inflammatory response in 6h of observation. The findings of this study
corroborated those of Dekker et al. [64] in which physical exercise did not
significantly change the IL-6 values, although a trend towards reduction was
perceived when compared with the control group.
In the study of Tyldum et al. [65] when they studied vasodilation
mediated by flow after lipid overload, with and without physical exercise,
showed evidence that with high intensity, periodized physical activity, the
vessel diameter increased when compared with that of the control, demonstrating
vasodilation secondary to physical activity.
Physical exercise has been associated with an increase in the nitric
oxide synthase enzyme, with an influence on the increase in nitric oxide,
providing a protective effect against endothelial dysfunction through physical
exercise. Physical exercise also induces the release of extracellular
superoxide dismutase which, according to Vuorimaa
[61], is an enzyme that acts in the antioxidant process.
Studies have shown evidence that dietary supplementation with
polyunsaturated fatty acids acted on reducing inflammatory response [66]. The
beneficial effects of dietary supplementation with polyunsaturated fatty acids
on exercise performance and on oxidative balance of physical activity have also
been shown in other studies [67] although the effects associated with intense
physical activity on the immune response are still contradictory [68].
According to Capó et al. [69], exercise
increased the activated levels of the anti-inflammatory response, increasing
anti-inflammatory gene expression after the exercise, particularly in the group
of young individuals. Recent studies have reinforced the idea that physical
exercise attenuates the inflammatory response [70].
Fatty
acids and oxidative stress
Oxidative stress formation, according to Ghisi
[56] would be one of the main factors responsible for triggering atherogenesis,
and that the superoxide anion (O2-) and oxidized LDL would be the
main free radicals involved in this process. In addition to this, Reactive
Oxygen Species (ROS) could interact with NO and form the peroxynitrite
anion (ONOO-) and nitrogen dioxide (NO2), which would be
responsible for potentializing the inflammatory lesion, favoring the
progression of atherosclerosis [61].
Lipid overload induces an increase in triglyceride-rich lipoprotein - TRLp, reduction in HDL and hyperinsulinemia. This metabolic
condition leads to the formation of free radicals, which are reduced, according
to the antioxidant capacity (endogenous and/or exogenous) present, determining
oxidative stress. The free radicals stimulate tissues to secrete cytokines
(TNF-α, IL-1 and IL-6), probably through the macrophages, thus stimulating
the formation of adhesion molecules. The generation of reactive oxygen species
diminishes the bioavailability of free NO, resulting in a lower level of
endothelium-dependent vasodilation, and also in the formation of peroxynitrite (ONOO-), a potent and long lasting oxidant. These processes are associated with
the genesis and progression of atherosclerotic lesions [60].
In the human body, there are lines of defense against the
atherosclerotic process, among them the antioxidant enzymes nitric oxide (NO)
and endothelial nitric oxide synthetase (eNOS);
superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX). NO
is an important factor responsible for the relaxation of arterial vessels:
depending on the medium in which this may function as an oxidant or reducer,
its oxidation produces nitrites and nitrates. Its anti-atherosclerotic
functions includes: inhibition of adhesion and
migration of leukocytes, impeding platelet aggregation; reduction in
endothelial permeability to lipoprotein macromolecules; impeding the
sub-endothelial accumulation of LDLc and its
oxidation, among others. These factors, among others, are related to modulation
of the inflammatory response [71].
According to the data from the study of Tyldum [65] the effects of a
diet rich in fats on reducing the antioxidant capacity appear, on an average,
30 minutes after ingestion.
Exercise
and oxidative stress
Light to moderate physical exercise performed regularly is recommended
for the maintenance of health and prevention of innumerable diseases It also
reduces the production of oxidants and occurrence of oxidative damage; improves
the antioxidant defense system; increases the resistance of organs and tissues
against the harmful action of the ROS, and diminishes systemic inflammation
[69]. However, there is a body of evidence suggesting that physical exercise,
particularly the more intense types, are associated with both muscular damage
and elevated ROS production [68].
In the study of Jong-Shyan et al. [72],
when they tested different intensities of physical exercise in sedentary
individuals with 40% VO2max, with 60% VO2max and with 80% VO2max
for 40 min, they verified that in the acute form, the highest intensity
resulted in higher production of oxidized LDL. This caused an increase in the
reactive oxygen species in the monocytes, when compared with the light and
moderate intensities. They concluded that in their study, acute, high intensity
physical exercise caused greater oxidative stress in sedentary individuals.
The study of Tyldum et al. [65] about the effect of acute
exercise of different intensities on the antioxidant capacity, showed that both
moderate and high intensity exercises interfered positively in the reduction of
antioxidant capacity influenced by diet, however, the results were more
expressive for the high intensity exercises.
According to Tyldum et al. [65] exercise and antioxidant function
revealed an interesting paradox; in the acute form, an increase occurs in the
levels of free radicals in the blood and muscle, which may be responsible for
the inactivity of large quantities of nitric oxide and negatively change the
endothelial impact mediated by vasodilation. However, the results of the study
revealed an increase in the antioxidant capacity in the acute stage of
exercise. The authors suggested that during the acute stage of the exercise, a
transfer of antioxidants occurs between the muscle and vasculature, favoring
the balance towards an inclination to favor the antioxidant effect, providing a
similar effect to that offered by a diet containing antioxidants.
Sureda et al.
[71] showed evidence that acute physical exercise performed at high intensities,
induced oxidative damage in the blood cells such as erythrocytes and
lymphocytes, but not in the neutrophils.
Habitual low to moderate intensity exercise is responsible for an
increased cellular antioxidant defense system; reduction in lipid peroxidation,
and protective effect against diseases associated with chronic inflammation
[69].
Some studies have pointed out that intense exercises are associated with
an increase in free radical formation. In other studies, acute and intense
exercises presented significant and rapid responses in endothelial function.
However, the intensity of exercise necessary to cause antioxidant responses is
still controversial in the literature.
Below, please find four summaries showing the articles presented in this
section, with reference to the effect of physical exercise on the inflammatory
response, endothelial dysfunction and oxidative stress.
Frame
3 - Effect of physical exercise on inflammatory
response, endothelial dysfunction and oxidative stress
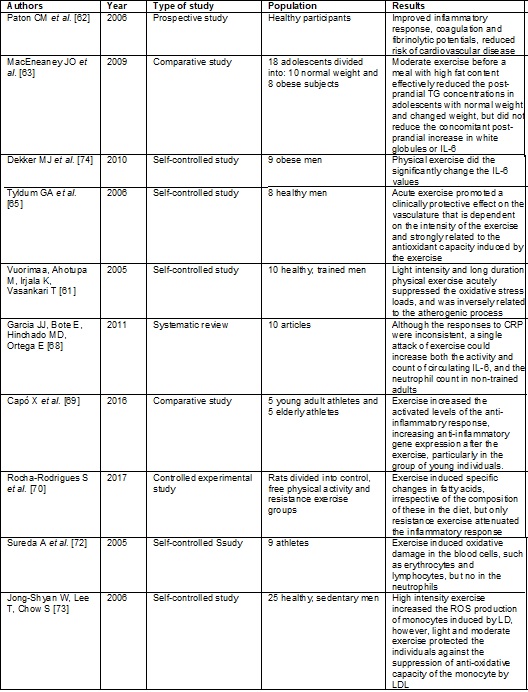
Conclusion
Fatty acids, depending on their biochemical characteristics and spatial
configuration, have a different effect on cardiovascular health, however,
studies still show contradictory results on the therapeutic use of certain
fatty acids. Physical exercise seems to benefit the metabolism of fatty acids
and mitigate complications secondary to the consumption of certain fatty acids,
in addition to enhancing the positive effects of different fatty acids.
However, variants of physical exercise, such as intensity, duration, time of
observation of the effects of the results, limit the authors to conclude, with
a certain degree of certainty, the effect of physical exercise on fatty acids
and secondary complications, since studies in the literature remain
contradictory.
Author contribution
Conception and research design: Wagmacker DS, Ladeia AMT. Data collection: Wagmacker DS, Oliveira AM, Oliveira EC, Santos ACN. Data analysis and interpretation:
Wagmacker DS, Oliveira AM, Santos ACN, Rodrigues LEA.
Writing of the manuscript: Wagmacker DS, Oliveira AM, Oliveira EC, Rodrigues LEA. Critical revision of the manuscript for important intellectual content: Ladeia AMT.
Academic
link
This article represents honest work and the validity of its results can
be certified. Furthermore, this article is part of Djeyne
Silveira Wagmacker M.Sc Thesis for the Bahian School of Medicine and
Public Health Post Graduate Course. And all authors declare no competing
interest. This work was supported by National Council for Scientific and
Technological Development (CNPQ).
Potential
conflict of interest
Has
no potential conflict of interest.
References
- Moran
AE, Forouzanfar MH, Roth GA, Mensah GA, Ezzati M, Murray CJ et al. Temporal trends in ischemic
heart disease mortality in 21 world regions, 1980 to 2010: the
Global Burden of Disease 2010 study. Circulation 2014;129:1483-92.
https://doi.org/10.1161/circulationaha.113.004042
- Finucane
MM, Stevens GA, Cowan MJ, Danaei G, Lin JK, Paciorek CJ et al. National, regional, and global trends in
body-mass index since 1980: systematic analysis of health examination surveys
and epidemiological studies with 960 country-years and 9.1 million
participants. Lancet 2011;377:557-67.
https://doi.org/10.1016/s0140-6736(10)62037-5
- Joint
ESC. Guidelines Atherosclerosis 2016;252:207-74.
- Gottlieb
MG, Bonardi G, Moriguchi
EH. Fisiopatologia
e aspectos inflamatórios da aterosclerose. Scientia
Medica 2005;15(3):203-7.
- Shivappa, N, Blair CK, Prizment AE, Jacobs DR, Steck SE,
Hébert JR. Association between inflammatory potential of diet and mortality in
the Iowa Women’s Health study. Eur J Nutr
2016;55(4):1491-502. https://doi.org/10.1007/s00394-015-0967
- Martin CA, Matshushita M, Souza NE. Ácidos graxos trans: implicações nutricionais e fontes na dieta. Rev Nutr 2004;17(3):351-9.
https://doi.org/10.1590/s1415-52732004000300009
- Costa AGV, Bressan J, Sabarense CM. Acidos
graxos trans: Alimentos e efeitos na saúde. Archivos
Latinoamericanos de Nutricion
2006;56(1):1-9.
- Oomen CM, Ocke
MC, Feskens EJ, Van Erp-Baart
MA, Kok FJ, Kromhout D.
Association between trans fatty acid intake and 10-year risk of coronary heart
disease in the Zutphen Elderly Study: a prospective
population-based study. Lancet 2001;357(1):746-51.
https://doi.org/10.1016/s0140-6736(00)04166-0
- Perez-Matos
MC, Morales-Alvarez MC, Mendivil CO. Lipids: a
suitable therapeutic target in diabetic neuropathy? J
Diabetes Res 2017;2017:6943851. https://doi.org/10.1155/2017/6943851
- Lopez-Garcia
E, Schulze MB, Meigs JB, Manson JE, Rifai N, Stampfer MJ et al. Consumption of trans fatty acids is related to plasma biomarkers of
inflammation and endothelial dysfunction. J Nutr
2005;135(1): 562-6.
- Jabbour G, Iancu
H-D, Paulin A, Lavoie J- M, Lemoine-Morel S, Zouhal
H. Effects of acute supramaximal cycle exercise on plasma ffa
concentration in obese adolescent boys. PLoS One
2015;10(6):e0129654.
https://doi.org/10.1371/journal.pone.0129654
- Bilet L, Brouwers B, van Ewijk PA, Hesselink MK, Kooi ME, Schrauwen P, et al. Acute exercise does not decrease liver
fat in men with overweight or NAFLD. Sci Rep 2015;13(5):9709.
https://doi.org/10.1038/srep09709
- Thivel D, Metz L,
Julien A, Morio B, Duche P.
Obese but not lean adolescents spontaneously decrease energy intake after
intensive exercise. Physiol Behav
2014;123:41-6.
https://doi.org/10.1016/j.physbeh.2013.09.018
- Bodel NG, Gillum T. 90 Minutes of moderate-intensity
exercise does not attenuate postprandial triglycerides in older adults. Int J Exerc Sci 2016;9(5):677-84.
- Davitt
PM, Arent SM, Tuazon MA, Golem DL, Henderson GC.
Postprandial triglyceride and free fatty acid metabolism in obese women after
either endurance or resistance exercise. J Appl Physiol
2013;114:1743-54.
https://doi.org/10.1152/japplphysiol.00095.2013
- Durstine JL, Thompson PD.
Exercise in the treatment of lipid disorders. Cardiol Clin
2001;19:471-88. https://doi.org/10.1016/s0733-8651(05)70230-7
- Rodrigues LEA.
Lipídios: aspectos bioquímicos e médicos. Salvador/Bahia: Ed UFBA; 2006.
- Mäkinen S, Nguyen
HY, Skrobuk P, Koistinen
HA. Palmitate and oleate exert differential effects on
insulin signalling and glucose uptake in human
skeletal muscle cells. Endocr Connect 2017;5:17-0039. https://doi.org/10.1530/EC-17-0039
- Kadhum AA, Shamma
MN. Edible lipids modification processes: A review. Crit
Rev Food Sci Nutr 2017;57(1):48-58.
https://doi.org/10.1080/10408398.2013.848834
- Moreira NX, Curi R,
Mancini Filho J. Acidos graxos: uma revisão. Nutrire 2002;24:105-23.
- Lima V. Cuidado com os
triglicerídeos! (bioquímica). In: Somos Físicos; 2016 [acesso em 2019 maio 31].
Disponível em:
http://www.vanialima.blog.br/2016/08/cuidado-com-o-triglicerides-bioquimica.html
- Brito
MS, Villavicencio ALCH, Mancini-Filho J. Effects of irradiation on trans fatty
acids formation in ground beef. Radiat Phys Chem
2002;63:337-40. https://doi.org/10.1016/s0969-806x(01)00647-8
- Aued-Pimentel
S et al. Ácidos
graxos saturados versus ácidos graxos trans em
biscoitos. Rev Inst Adolfo Lutz 2003;62(2):131-7.
- Sugano
M, Ikeda I. Metabolic interactions between essential and trans fatty acids. Curr Opin Lipidol
1996;7:38-42.
https://doi.org/10.1097/00041433-199602000-00009
- Abreu
P, Leal-Cardoso JH, Ceccatto VM, Hirabara
SM. Regulation of muscle plasticity and trophism by fatty acids: A short
review. Rev Assoc Med Bras 2017;63(2):148-55.
https://doi.org/10.1590/1806-9282.63.02.148
- Sandres TA, Oakley FR, Crook
D, Coper JA, Miller GJ. High intakes of trans monounsaturated fatty acids taken
for 2 weeks do not influence procoagulant and fibrinolytic risk markers for CHD
in young healthy men. Br J Nutr 2003;89(6):767-76.
https://doi.org/10.1079/bjn2003850
- Perez-Matos
MC, Morales-Alvarez MC, Mendivil CO, Lipids: a
suitable therapeutic target in diabetic neuropathy? J Diabetes Res 2017.
https://doi.org/10.1155/2017/6943851
- Zapolska-Downar D, Kosmider
A, Naruszewicz M. Trans fatty acids induce apoptosis
in human endothelial cells. J Physiol Pharmacol
2005;56(4):611-25.
- Sun
Q, Ma J, Campos H, Hankinson SE, Manson JE, Stampfer MJ et al. A prospective study of trans fatty acids in erythrocytes and risk of
coronary heart disease. Circulation 2007;115(14):1858-65.
https://doi.org/10.1161/circulationaha.106.679985
- Chajès V, Biessy
C, Ferrari P, Romieu I, Freisling
H, Huybrechts I et al. Plasma elaidic acid level as
biomarker of industrial trans fatty acids and risk of weight change: Report
from the EPIC Study. PLoSOne 2015;10(2):e0118206. https://doi.org/10.1371/journal.pone.0118206
- Ford
PA et al. Trans fatty acid intake is related to emotional affect
in the Adventist Health Study-2. Nutr Res
2016;36(6):509-17. https://doi.org/10.1016/j.nutres.2016.01.005
- Sposito
AC et al. IV
Diretriz Brasileira sobre Dislipidemias e Prevenção da Aterosclerose:
Departamento de Aterosclerose da Sociedade Brasileira de Cardiologia. Arq Bras Cardiol 2007;88(1)1 2-19.
- Abbott
SK, Else PL, Hulbert AJ. Membrane fatty acid composition of rat skeletal muscle
is most responsive to the balance of dietary n-3 and n-6 PUFA. Br J Nutr 2010;103(4):522-9.
https://doi.org/10.1017/s0007114509992133
- Veliça PP, Khanim FL, Bunce CM. Prostaglandin D2 inhibits C2C12 myogenesis. Mol Cell Endocrinol
2010;319(1-2):71-8. https://doi.org/10.1016/j.mce.2010.01.023
- Fostok SF, Ezzeddine
RA, Homaidan FR et al. Interleukin-6 and
Cyclooxygenase-2 downregulation by fatty-acid fractions of Ranunculus constantinopolitanus. BMC Complementary and Alternative
Medicine 2009;9:44. https://doi.org/10.1201/b16611-5
- Hamley S. The effect of replacing
saturated fat with mostly n-6 polyunsaturated fat on coronary heart disease: a
meta-analysis of randomised controlled trials. Nutr J 2017;16(30).
https://doi.org/10.1186/s12937-017-0254-5
- Kazantzis M, Stahl A. Fatty acid
transport proteins, implications in physiology and disease. Biochimica et Biophysica
Acta 2012;1821(5):852-7. https://doi.org/10.1016/j.bbalip.2011.09.010
- Aon MA, Bhatt N, Cortassa SC. Mitochondrial and cellular mechanisms for managing lipid excess.
Frontiers in Physiology 2014;5(282).
https://doi.org/10.3389/fphys.2014.00282
- Sztalryd C, Kimmel AR.
Perilipins: lipid droplet coat proteins adapted for tissue-specific energy
storage and utilization, and lipid cytoprotection. Biochimie 2014;96:96-101.
https://doi.org/10.1016/j.biochi.2013.08.026
- Morales
PE, Bucarey JL, Espinosa A. Muscle lipid metabolism:
role of lipid droplets and perilipins. Journal of Diabetes Research
2017:1789395. https://doi.org/10.1155/2017/1789395
- Ewa H, Agnieszka K, Tomasz S, Adrian
C. The role of fatty-acid transport proteins (FAT/CD36, FABPpm,
FATP) in lipid metabolism in skeletal muscles. Postepy
Hig Med Dosw 2008;62:433-41.
- Smith
BK, Bonen A, Holloway GP. A dual mechanism of action
for skeletal muscle FAT/CD36 during exercise. Exercise and Sport Sciences
Reviews 2012;40(4):211-7. https://doi.org/10.1097/jes.0b013e31825eb263
- Jayewardene
AF, Mavros Y, Reeves A, Hancock DP, Gwinn T, Rooney
KB. Interactions between fatty acid transport proteins, genes that encode for
them, and exercise: a systematic review. J Cell Physiol
2016;231(8):1671-87. https://doi.org/10.1002/jcp.25281
- Watt
MJ, Cheng Y. Triglyceride metabolism in exercising muscle. Biochim
Biophys Acta 2017;23(17):30119-1.
https://doi.org/10.1016/j.bbalip.2017.06.015
- Badin
PM, Loubiere C, Coonen M et
al. Regulation of skeletal muscle lipolysis and oxidative metabolism by the
co-lipase CGI-58. Journal of Lipid Research 2012;53(5):839-48.
https://doi.org/10.1194/jlr.m019182
- Turnbull
PC, Longo AB, Ramos SV, Roy BD, Ward WE, Peters SJ. Increases in skeletal
muscle ATGL and its inhibitor G0S2 following 8 weeks of endurance training in
metabolically different rat skeletal muscles. American Journal of Physiology.
Regulatory, Integrative and Comparative Physiology 2016;310(2):R125–R133.
https://doi.org/10.1152/ajpregu.00062.2015
- Roepstorff
C, Vistisen B, Donsmark M et al. Regulation of
hormone-sensitive lipase activity and Ser563 and Ser565 phosphorylation in
human skeletal muscle during exercise. J Physiol
2004;560(2):551-62. https://doi.org/10.1113/jphysiol.2004.066480
- Casas
M, Figueroa R, Jorquera G, Escobar M, Molgó J, Jaimovich E. IP(3)-dependent,
post-tetanic calcium transients induced by electrostimulation of adult skeletal
muscle fibers. The Journal of General Physiology 2010;136(4):455-67.
https://doi.org/10.1085/jgp.200910397
- Barres
R, Yan J, Egan B, Treebak JT, Rasmussen M, Fritz T et
al. Acute exercise remodels promoter methylation in human skeletal muscle. Cell
Metab 2012;15:405-11.
https://doi.org/10.1016/j.cmet.2012.01.001
- Egan
B, Zierath JR. Exercise metabolism and the molecular
regulation of skeletal muscle adaptation. Cell Metab 2013;17:162-84. https://doi.org/10.1016/j.cmet.2012.12.012
- Kim
J, Lee KP, Lee DW, Lim K. Piperine enhances
carbohydrate/fat metabolism in skeletal muscle during acute exercise in mice.
Nutrition & Metabolism 2017;14:43.
https://doi.org/10.1186/s12986-017-0194-2
- Homer
AR, Fenemor SP, Perry TL, Rehrer
NJ, Cameron CM, Skeaff CM et al. Regular activity
breaks combined with physical activity improve postprandial plasma
triglyceride, nonesterified fatty acid, and insulin
responses in healthy, normal weight adults: A randomized crossover trial. J
Clin Lipidol 2017;19(17):30351-3.
https://doi.org/10.1016/j.jacl.2017.06.007
- Petridou A, Chatzinikolaou
A, Avloniti A, Jamurtas A, Loules G, Papassotiriou I et al.
Increased triacylglycerol lipase activity in adipose tissue of lean and obese
men during endurance exercise. J Clin Endocrinol Metab
2017;12:3945-52.
https://doi.org/10.1210/jc.2017-00168
- Nojiri S, Daida
H. Atherosclerotic cardiovascular risk in Japan. Japanese Clinical Medicine
2017;8; 117906601771271. https://doi.org/10.1177/1179066017712713
- Iso
H, Cui R, Date C et al. C-reactive protein levels and risk of mortality from
cardiovascular disease in Japanese: the JACC Study. Atherosclerosis 2009;207:291-7.
https://doi.org/10.1016/j.atherosclerosis.2009.04.020
- Ghisi GLM et al. Exercício
físico e disfunção endotelial. Arq Bras Cardiol 2010; 95(5):130-7.
https://doi.org/10.1590/s0066-782x2010001500025
- McClean
CM, McLaughlin J, Burke G, Murphy MH, Trinick T, Duly
E, Davison GW. The effect of acute aerobic exercise on pulse wave velocity and
oxidative stress following postprandial hypertriglyceridemia in health men. Eur
J Appl Physiol 2007;100:225-34.
https://doi.org/10.1007/s00421-007-0422-y
- Stapleton
PA, Goodwill AG, James ME. Hypercholesterolemia and microvascular dysfunction:
interventional strategies. J Inflamm 2010;18;7:54. https://doi.org/10.1186/1476-9255-7-54
- Nappo F, Esposito K, Cioffi M, Giugliano G, Molinari AM, Paolisso
G et al. Postprandial endothelial activation in healthy subjects and in type 2
diabetic patients: role of fat and carbohydrate meals. J Am
Coll Cardiol 2002;39:1145-50.
https://doi.org/10.1016/s0735-1097(02)01741-2
- Pinho, RA. Araújo MC, Ghisi GLM et al. Doença arterial coronariana, exercício
físico e estresse oxidativo. Arq Bras Cardiol 2010;94(4). https://doi.org/10.1590/s0066-782x2010000400018
- Vuorimaa T, Ahotupa
M, Irjala K, Vasankari T.
Acute prolonged exercise reduces moderately oxidized LDL in healthy men. Int J Sports Med 2005;26:420-5.
https://doi.org/10.1055/s-2004-821142
- Paton
CM et al. Hemostatic response to postprandial lipemia before and
after exercise training. J Appl Physiol 2006;101:316-21.
https://doi.org/10.1152/japplphysiol.01363.2005
- MacEneaney JO et al. effect of
prior exercise on postprandial lipemia and markers of inflammation and endothelial
activation in normal weight and overweight adolescent boys. Eur J Appl Physiol 2009; 106:721-9.
https://doi.org/10.1007/s00421-009-1073-y
- Dekker
MJ, Graham ET, Ooi TC, Robinson LE. Exercise prior
to fat ingestion lowers fasting and postprandial VLDL and decreases adipose
tissue IL-6 and GIP receptor mRNA in hypertriacylglycerolemic
men. J Nutr Biochem 2010;21:983-90.
https://doi.org/10.1016/j.jnutbio.2009.08.004
- Tyldum
GA et al. Endothelial dysfunction induced by postprandial
lipemia: complete protection afforded by high intensity aerobic interval
exercise. J
Am Coll Cardiol.
2009;53(2):200-2006.
- Medina-Remon A, Tresserra-Rimbau A, Pons
A, Tur JA, Martorell M, Ros E et al. Effects of total
dietary polyphenols on plasma nitric oxide and blood pressure in a high
cardiovascular risk cohort. The predimed randomized
trial. Nutr Metab
Cardiovasc Dis 2015;25:60-7.
https://doi.org/10.1016/j.numecd.2014.09.001
- Mickleborough T.D. Omega-3
polyunsaturated fatty acids in physical performance optimization. Int J Sport Nutr Exerc Metab 2013;23:83-96.
https://doi.org/10.1123/ijsnem.23.1.83
- Garcia
JJ, Bote E, Hinchado MD,
Ortega E. A single session of intense exercise improves the inflammatory
response in healthy sedentary women. J Physiol Biochem 2011;67:87-94. https://doi.org/10.1007/s13105-010-0052-4
- Capó X, Martorell M, Sureda A, Riera J, Drobnic F, Tur, JA, Pons A. Effects of almond- and olive
oil-based docosahexaenoic- and vitamin e-enriched beverage dietary
supplementation on inflammation associated to exercise and age. Nutrients 2016;8(10):619.
https://doi.org/10.3390/nu8100619
- Rocha-Rodrigues S,
Rodríguez A, Gonçalves IO, Moreira A, Maciel E, Santos S et al. Impact of physical exercise on visceral adipose tissue fatty acid
profile and inflammation in response to a high-fat
diet regimen. Int J Biochem Cell Biol 2017;87:114-24.
https://doi.org/10.1016/j.biocel.2017.04.008
- Sureda A, Tauler
P, Aguiló A, Cases N, Fuentespina
E, Córdova A et al. Relation
between oxidative stress markers and antioxidant endogenous defenses during
exhaustive exercise. Free Radic Res
2005;39(12):1317-24. https://doi.org/10.1080/10715760500177500
- Jong-Shyan W, Lee T, Chow S. Role of exercise intensities in
oxidized low-density lipoprotein-mediated redox status of monocyte in men. J Appl
Physiol 2006;101(3):740-4.
https://doi.org/10.1152/japplphysiol.00144.2006